The purpose of this post is to very briefly describe how interlaminar astrocytes and Bergman glia form perpendicular cylindrical arrays around columns of neurons.
This post is part two of a three part series describing the possible function of quasi static magnetic fields formed by cylindrical arrays of perpendicular astrocytes. The next post looks at how cylindrical arrays of perpendicular astrocytes might form a magnetic shield or magnetic cloak.
Background
Micro-columns are columnar groupings of neurons and astrocytes that form repeating structures in some parts of the neocortex.
Micro-columns were first described by Mountcastle (1957) as “neurons of the somatic cortex [that] are organized into vertical columns or cylinders which extend perpendicular: through the cortical layers.”

Diagram (DeFelipe, J. 2015) representing a neocortical micro column, based mainly around the layer 5 pyramidal neurons.
Micro-columns are assumed to commonly map between an internal topology and an external topology (Tommerdahl 2005). In that sense cortical columns are semantic structures much like cortical maps have a semantic basis. In some cortical maps they confirm an evolutionary or functional advantage and may have evolved or develop.
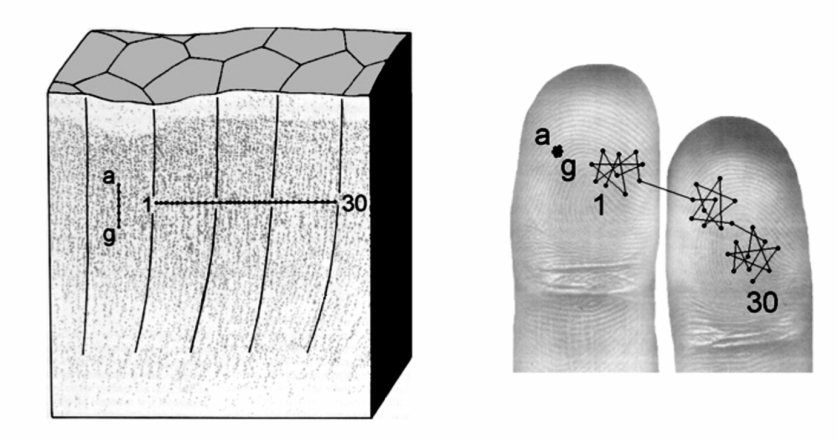
Diagram (Tommerdahl 2005) showing how the micro columns in the somatosensory cortex map to sensory finger nerves.
Micro-columns are often based on pyramidal neurons whilst, excitatory spiny stellate cells and inhibitory double-bouquet cells, excite and inhibit micro-columns in adjacent micro-columns (Tommerdahl 2005).

Diagram (Tommerdahl 2005) showing how the micro columns excite and inhibit adjacent micro-columns.
Only some parts of the cortex contain repeating micro-column structures and their detailed structure and function varies between mammalian species and even individuals (Jones 2000). Micro-columns are more prevalent in the sensory cortex, such as visual, temporal and striate cortex. These regions correlate with the more laminated areas in cortex proposed by (John 2021).
There is a continuous debate about the function of micro-columns (Buxhoeveden 2005). A canonical microcircuit that corresponds to cortical columns has not been found across the cortex and the size of their structure could be the result of a common evolutionary journey (Horton 2005).
The size and existence of micro-columns is dependent on the individual function of the map in which they occur. There is a spectrum in the literature between larger columnar arrays such as the barrel and ocular dominance columns and smaller micro-columns and cortical maps with no micro-columns. Some columns are 600um to 900um across whilst others are 30um-80um across (Jones 2000). These varying sizes of columns have columns have been given names such as macro, mini or micro columns.
Distribution of Astrocytes in Micro-columns
Astrocyte morphology is strongly confined within cortical micro-column boundaries in mammals, ranging from mice to human, as measured by the lower density of astrocytes at boundaries of micro-columns (Eilam 2016). This distribution was found for cytoplasmic astrocytes in the mouse barrel cortex and rat auditory rat cortex and for the interlaminar astrocytes in the human colour-blobs cortex and human striate cortex and V1 of the human visual cortex.

Picture (Eilam 2016) showing the distribution of astrocytes in the layer 5 of the mouse barrel cortex.
Astrocytic Calcium
The protoplasmic (not interlaminar) astrocytic calcium response to visual stimulus was found to vary between mammalian species and was dependent on having cortical micro columns. It was found (López-Hidalgo 2017) that a species with cortical V1 orientation micro-columns (ferrets) has sustained calcium response to visual stimulus, whereas species with no cortical V1 orientation micro-columns (mice) had weak calcium response to visual stimulus. In ferrets astrocytes are twice as big as mice, and overlap 50% of its territory with 6-8 neighboring astrocytes unlike mice.

Diagram showing neural and astrocytic calcium response to ferret (left) and mouse (right).
In contrast, the mouse cortical A1 auditory (tonal) micro-columns were found to have a large sustained calcium response to stimulation (Tischbirek 2019). The mouse barrel cortex is also known to have a micro-column structure and also has sustained calcium response.
Magnetic Fields in Astrocytes
Magnetic fields can travel through tissue similarly to free space as tissue has the same magnetic permeability (magnetic) as free space (Caruso 2017). However electric fields are limited in their ability to pass through cellular tissue and bulk water. Previous posts have shown that confined water inside neurons can orientate water so that the permittivity (electric) is greatly reduced along the line of the cell, but not directly through the cell wall.
It was hypothesised that owing to the long, thin, geometry of intralaminar astrocytes (and possibly Bergman glia) they can produce bio-electric and and bio-magnetic fields, which can “coherently integrate external and internal signals between neurons and astrocytes by ephaptic interaction”. (Martinez-Banaclocha M. 2018).
Martinez-Banaclocha (2018) speculated that the magnetic fields produced by astrocytes creates ephaptic coupling. Ephaptic coupling refers to interaction between neurons and astrocytes through the electric and magnetic fields in the extra cellular field. Ephaptic coupling has generally been used to refer to coupling between action potentials along axons, but it also has a small effect on dendrites where it effects post synaptic potentials.
Diagram (Martinez-Banaclocha 2018) showing how a moving Ca+2 charge induces a magnetic field inside an astrocyte.

Ca+2 moves continuously inside astrocytes during cycles of action potentials.

Diagram (Gai 2020) showing a continuous movement of Ca+2 during cycles of action potentials.
Magnetic Fields in Micro-columns
Martinez-Banaclocha (2020) further hypothesised that astrocytes surrounding cortical micro-columns produce magnetic fields which ephaptic couple with astrocytes and neurons in adjacent micro-columns. This hypothesis suggests there is a ephaptic coupling between microcolumns.

Diagram (Martinez-Banaclocha M. 2020) showing magnetic and electric fields in microcolumns.
However the previous study (Eilam 2016) showed that astrocytes do not form a boundary around micro-columns (see image above). In fact astrocytes are spread evenly within the micro-column and are reduced at the boundary. This means the micro-column structure surrounded by astrocytes suggested Martinez-Banaclocha M. (2020) may not be correct.
An alternatively biologically realistic model, that retains the concept of vertical cylindrical arrays of astrocytes and the movement of Ca+2 molecules, has been suggested by Colombo (2002). In this model cylindrical arrays of astrocytes containing moving Ca+2 ions expands outwards concentrically.
It was found that the “sphere of influence” of diffusing Ca+2 molecules may control a radial arrangement of interlaminar processes (Colombo 2002). This model suggests that interlaminar astrocytes trigger adjacent astrocytes through electrical synapses, to form an efficient, radial cylindrical structure containing moving Ca+2. This moving Ca+2 creates magnetic fields.

Diagram (Colombo 2002) showing the model of a cross section of the expansion of a Ca+2 wave across interlaminar astrocytes in the horizontal axis, where each hexagon is a micro-column.
A similar radial spread of Ca+2 though vertical Bergman glia (astrocytes) has also been found in the cerebellum (Hooglanda 2009). This suggests that both interlaminar astrocytes and Bergman glia form vertical arrays of moving Ca+2 in both the cortex and cerebellum.

Diagram (Hooglanda 2009) showing the cross section of the expansion of a Ca+2 wave across cerebellum, in the horizontal axis.
Next Step
The next post will show the possible function of cylindrical magnetic fields produced by astrocytes in the cortex and glia in the cerebellum.
